Innervated Myotendinous Cylinders Alterations in Human Extraocular Muscles in Patients With Strabismus
Article information
Abstract
Purpose
To analyze innervated myotendinous cylinders (IMCs) in the extraocular muscles (EOMs) of normal subjects and strabismic patients.
Methods
The rectus muscles of 37 subjects were analyzed. Distal myotendinous specimens were obtained from 3 normal subjects, 20 patients with acquired strabismus, 11 with infantile strabismus, and from 3 with congenital nystagmus, and were studied by using light microscopy. Some specimens (6 rectus muscles) were also examined by transmission electron microscopy.
Results
IMCs were found in the distal myotendinous regions of EOMs. The IMCs of patients with acquired strabismus showed no significant morphological alterations. However, significant IMCs alterations were observed at the distal myotendinous junction of patients with congenital strabismus and congenital nystagmus.
Conclusions
This study supports the notion that IMCs in human EOMs function mainly as proprioceptors, along with effector properties, and a disturbance of ocular proprioceptors plays an important role in the pathogenesis of oculomotor disorder. We suggest that a proprioceptive feedback system should be stimulated and calibrated early in life for the development of binocular vision.
Proprioceptive input from EOMs is supposed to be important for the control of ocular alignment and for the development of binocular vision.1-4 Although the role of ocular proprioception on the pathogenesis of heterotropia is unknown, several authors have attributed some oculomotor disorders to proprioceptive alterations.5-7
In mammalian limb muscles, the classic proprioceptors are muscle spindles and Golgi tendon organs. However, the proprioceptive component of EOMs differs from these of other skeletal muscles and shows striking interspecies variations. In EOMs, muscle spindles are found in humans and two-toed ungulates,8 but a detailed structural analysis of human adult and infant EOMs casts doubt on their viability as proprioceptors.9,10 Moreover, Golgi tendon organs have been widely reported to be absent in human EOMs.9,11-14
The principal proprioceptors of human EOMs are the so-called palisade endings. Dolgiel15 first reported that the commonest tendon nerve ending in EOMs is composed of myelinated nerve fibers that penetrate the tendon, and then turn back to divide into several approximately parallel running branches investing the tip of single muscle fibers.
These are called palisade endings and have been described in several animals. The structure and distribution of these endings were confirmed in human EOMs by Richmond et al.13 Nervous end organs containing palisade endings are enclosed by a loose capsule of connective tissue cells and consist of a terminal portion of a single multiply innervated muscle fiber of the global layer and its attached tendon. This nervous end organ is referred to as an "innervated myotendinous cylinder" (IMC).16 Moreover, these properties distinguish IMCs from classic Golgi tendon organs.
It has been demonstrated that the EOMs of strabismic patients have normal motor nerve endings, mainly located in the center of EOMs.17-19 In contrast, ultrastructural data on the distal myotendinous junction of the EOMs in congenital strabismus indicates the presence of an altered proprioceptive innervation.7,20 This morphologic data supports the hypothesis that a disturbance of ocular proprioception, in the myotendinous junction, may play a role in the pathogenesis of oculomotor disorder. However, data on possible morphologic ocular proprioceptors alterations in various forms of strabismus is inadequate.
This study was undertaken to investigate the morphological features of ocular proprioceptors located at the distal myotendinous junction of non-infantile and infantile strabismus subjects and of congenital nystagmus subjects, and to compare these with those of normal subjects.
Materials and Methods
The light microscopic study was performed on 40 human horizontal rectus muscles obtained from 3 normal subjects and 34 patients with oculomotor disorders. Six control horizontal rectus muscles were obtained from the right globe of 3 multiorgan donors (2 muscles from each) a few hours after postmortem in conformity with legal requirements. The normal subjects had no disturbance of the oculomotor system. Patients with oculomotor disorders were composed of 20 patients with acquired strabismus, 11 with infantile strabismus, and 3 with congenital nystagmus. One horizontal rectus muscle from each patient with oculomotor disorder was obtained during resection of a rectus muscle.
The electron microscopic examination was carried out on 6 horizontal rectus muscles from the left globe of 2 multiorgan donors (8 month infant and 61-year-old female) and 4 patients, including 10-year-old subject with intermittent exotropia, 3-year-old with infantile exotropia, 49-year-old with acquired paralytic strabismus and 4-year-old with congenital nystagmus.
Methods for securing human tissues were humane, included proper consent and approval, and complied with the tenets of the Declaration of Helsinki and Austrian federal transplantation law.
The 20 patients with acquired strabismus consisted of 6 with paralytic strabismus, 5 with constant exotropia, 5 with intermittent exotropia, 3 with sensory strabismus, and 1 with secondary esotropia after retinal reattachment. The 11 patients with infantile strabismus consisted of 5 with infantile exotropia, 5 with infantile esotropia, and 1 with congenital third cranial nerve paralysis. Three normal subjects were 8 months, 52 and 61 years old respectively. The mean age in patients was 29.4 (range, 2-64) years in patients with acquired strabismus, 3.45 (range, 1-10) years in patients with infantile strabismus, and 4.3 (range, 3-6) years in patients with congenital nystagmus (Table 1).

Characters of subjects
Before surgery, all patients affected by strabismus demonstrated large-angle deviations (≥ 50 prism diopters), and all patients with congenital nystagmus showed large-angle face turn (≥ 30 degrees). All samples from the EOMs were obtained by excising a full width strip of rectus muscle during strabismus surgery. EOMs were placed on slight stretch and care was taken to include the distal myotendinous junction in the 8-12 mm long tissue strip.
Light Microscopy
All specimens were gently stretched with 2 forceps and fixed with pins to the bar during the fixation process to prevent folding and curling. The specimens were fixed for at least 24 hours in 10% buffered formalin and then embedded in paraffin for routine histopathological processing. Complete series of paraffin cross sections (10 µm) were cut on a microtome and mounted on glass slides. Paraffin cross sections were stained with hematoxylin and eosin at approximately 200 µm intervals, and we examined samples progressing from muscle to tendon to select the most distal section just before reaching the first evidence of tendon. From the distal part from this selected section, we made 5 serial sections at intervals of 50 µm, which totaled to 200 µm. Series of these 5 sections in the myotendinous junction were stained with hematoxylin and eosin. Total IMC counts in each EOM were evaluated based on an examination of these 5 serial sections with 50 µm intervals in the distal myotendinous junction (200 µm). When the IMC was found, 2 nearest sections (10 µm intervals) were impregnated with silver for light microscopy.
Electron Microscopy
For conventional transmission electron microscopy, tissues were immersed in a fixative solution consisting of 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.4) and allowed to remain in this solution for more than 12 hours. After rinsing in the same buffer, samples were postfixed in 1% osmium tetroxide in phosphate buffer 0.1 M (pH 7.4), dehydrated in ethanol at increasing concentrations, and embedded in epon. Serial semi-thin cross sections (40 µm) were examined. When the IMC was found, approximately 10 ultra-thin sections alternating with semi-thin ones were cut at appropriate intervals (50-60 nm), mounted on copper grids, stained with uranyl acetate and lead citrate, and examined under a Hitachi® H-7100 transmission electron microscope (Hitachi, Tokyo, Japan).
Results
Morphology of Innervated Myotendinous Cylinders
Light Microscopy
Resected tissue of EOMs were included part of a distal myotendinous junction. Small nerves enclosed by a loose capsule of connective tissue cells terminated in the myotendinous region.
The IMCs were identified with a light microscope by two investigators (SEP, HSS). Both observers were able to obtain completely concordant data from identical pieces of tissue. Consecutively, the number of IMCs was analyzed in all resected EOMs. Numerous IMCs were observed in all EOMs in the distal myotendinous regions. Golgi tendon organs or muscle spindles were not observed in any stained EOM region.
In the control EOMs, nerve ending and muscle fibers were sheeted by thin capsules, which constituted the IMCs (Fig. 1A). The presence of nerve fibers within the IMCs was revealed by silver staining (Fig. 1B). Examinations of the distal myotendinous junctions of EOMs of patients with acquired strabismus showed normal features. In the congenital strabismus EOMs, nerve endings were smaller in size than those of controls (Fig. 1C).

The cross section of the control medial rectus muscle at the distal myotendinous junction (1-A,B) and the cross section of medial rectus muscle of infantile intermittent exotropia. (1-C) (A) Innervated myotendinous cylinder (IMC) (arrow) is shown among the muscle fibers. The tip of muscle fiber (arrowheads) with a nerve ending is encapsulated by a loose connective tissue (C), and those compose a normal IMC. H & E stain. Scale bar, 100 µm. (B) Nerve ending (arrow) is shown in IMC. Silver stain. Scale bar, 100 µm. (C) The small fragmented nerve endings (arrow) is seen. H & E stain. Scale bar, 50 µm.
The IMCs passed forward from distal muscle towards tendon. Many of them continued into tendon for a short distance, approximately 100 µm. The distance of neighboring nerve fibers increased gradually from distal to proximal muscle fibers (Fig. 2).

Cross sections of a muscle compartment of a myotendinous junction the control medial rectus muscle taken at intervals of approximately 50 µm from distal to proximal direction. Two innervated myotendinous cylinders (arrows) are shown close together at the distal part (A), but getting far from each other at the proximal part (C). H & E stain. Scale bar, 100 µm.
Electron Microscopy
At the distal myotendinous junction of control EOMs, several IMCs were observed near the muscle fibers. Within the IMC capsule, a single myelinated nerve fiber ran without branching, and the axon was completely sheathed by a Schwann cell. The IMC capsule also contained muscle fibers and fibrocytes (Fig. 3).

Ultrathin cross section through an innervated myotendinous cylinder (IMC) at the myotendinous junction of the control medial rectus muscle. (A) Muscle compartment of an IMC. The IMC is encircled by a capsule (C) of fibrocytes. A myelinated nerve (N) in IMC has already passed through the capsule and is completely surrounded by a Schwann cell (S). Inside the capsule, a muscle fiber (MF) and fibroblast (F) are also present. Scale bar, 5 µm. (B) Detailed image of the nerve terminal contacting the muscle fiber. Myelinated nerve fiber (M) contains mitochondria (arrowhead), rough endoplasmic reticulum (rER) and neurotubule (N). The synaptic cleft (arrow) is free of the basal lamina. Scale bar, 5 µm. (C) Tendon compartment of an IMC lying among collagen fibrils. It contains mitochondria (arrow) and clear vesicles (arrowhead). Scale bar, 2 µm.
The distal myotendinous junction in patients with acquired strabismus showed no significant morphological alterations. Most IMCs possessed normal features, however some Schwann cell degenerations were observed in the EOMs of patient with intermittent exotropia (Fig. 4). Examination also found atrophic muscle fiber and flattened Schwann cell process in some of the IMCs of patient affected by paralytic strabismus (Fig. 5).

Ultrathin cross section through an innervated myotendinous cylinder of medial rectus muscle in patient with intermittent exotropia. A nerve terminal is surrounded by a Schwann cell (S). Note flattened cytoplasmic process and focally concentric arrangement (arrow) of Schwann cell process. Muscle fiber (MF); Myelinated nerve fiber (M); Unmyelinated nerve fiber (UM). Scale bar, 5 µm.

Ultrathin cross section through an innervated myotendinous cylinder of lateral rectus muscle in patients with paralytic strabismus. (A) The IMC is encapsulated by a loose connective tissue (C). Inside the capsule, muscle fiber (MF) looks atrophic, and Z-disc (arrow) shows disoriented and irregular features. Myelinated nerve fiber (arrowhead). Scale bar, 5 µm. (B) Detailed image of the nerve terminal contacting the muscle fiber (MF). The nerve axon (A) looks normal, but cytoplasmic process of a Schwann cell (S) is flattened (arrow). Scale bar, 5 µm.
Most nerve fibers at the distal myotendinous junction in patient with infantile strabismus were observed to be altered. These modifications consisted of Schwann cell degeneration and axon alteration with maintenance of a practically normal general architecture (Fig. 6).
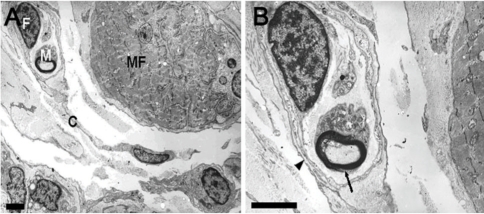
Ultrathin cross section through an innervated myotendinous cylinder of medial rectus muscle in patient with infantile intermittent exotropia. (A) Muscle fiber (MF) and myelinated nerve fiber (M) are surrounded by a capsule (C) of fibrocytes (F). The capsular investment is discontinuous. Scale bar, 5 µm. (B) Detailed image of the nerve terminal. In an uneven myelin sheath (arrow) abnormally enlarged axon containing innumerable neurofilaments is present. The basal lamina of Schwann cell process (arrowhead) is flattened and focally reduplicated. Scale bar, 5 µm.
Examinations of the distal myotendinous junction of patient with congenital nystagmus revealed small, anomalous nerve terminal endings. Some Schwann cells and axons lacked neurotubules and neurofilaments, and Golgi apparatuses and mitochondria were also absent. Axonal myelin was anomalous, discontinuous, and unevenly distributed, and unmyelinated nerve fibers demonstrated fragmented neurilemma, swelling, and degeneration. Some IMCs were devoid of nerve component (Fig. 7).

Ultrathin cross section through an innervated myotendinous cylinder of lateral rectus muscle in a patient with congenital nystagmus. Nerve endings (N) show empty space lacking of nerve component. Scale bar, 5 µm.
Discussion
Although still under debate, proprioceptive input from the EOMs probably plays a role in the control of ocular alignment. Steinbach and Smith5 have given supports for a proprioceptive contribution to spatial localization from the study of strabismus patients. They have demonstrated patients undergoing strabismus surgery for a second time showed more errors in perceiving shifts of eye position when compared with those undergoing their first surgery, and suggested that some nervous organs may be important for eye position proprioception. Palisading nerve endings in IMCs have been founded by Richmond et al.13 as the proprioceptive organs. Ruskell denied not only the existence of IMCs at birth in human,21 but also their role in proprioception in his review article.22 However, Lukas et al.23 have given important arguments against Ruskell's study, and suggested that IMCs may serve as proprioceptors as well as effectors.
Despite many studies on the subject, uncertainty persists of the exact roles of IMCs in visual spatial perception and ocular motor behavior. Several groups have suggested that IMCs are sensory,16,21,22,24-29 but others have considered them as motor30-32 structures, or both.23
Billig et al.26 identified sensory nerve endings in cat extraocular muscles by injecting neuronal tracers into the trigeminal ganglion, which contains the sensory cells that innervate the eye muscles. After the application, three types of nerve endings were labeled within EOMs, one type confirmed palisading endings of IMCs at the myotendinous junction of each EOM, whilst the other types were sensory terminals within the muscle belly. These results support that IMCs have a sensory function.
Lukas et al.23 investigated the anatomical structures of IMCs in human EOMs. They reported IMCs establish contacts with tendon fibrils and attach muscle fibers, and reveal sensorylike nerve terminals. However, they also presented the morphological features of motor terminals, which were confirmed by α-bungarotoxin staining. Based on these features, they proposed that IMCs might function as "propriocept-effectors", by combining sensory and motor qualities.
Another view have highlighted IMCs as motor structures and suggested that visual spatial perception might be served by efference copy.14,30-32 Sas and Scháb30 observed degenerated palisade endings in EOMs after removing small stereotactic lesions in oculomotor nuclei. Blumer et al.31 studied IMCs in rabbit EOMs, and based on fine structure and α-bungarotoxin binding suggested that myoneural contacts in IMCs are exclusively motor. A recent study on cat EOMs also provided evidence that palisade endings are exclusively motor. Specifically, it was found that palisade endings arise from nerve fibers that establish motor contacts on muscle fibers, and also that neuromuscular contacts in palisade endings exhibit α-bungarotoxin staining.32
Specific research attention has also been focused on the possible correlation between oculomotor disorders and structural modifications of proprioceptors at the myotendinous junction. However, data on the possible morphological alterations of IMCs in the various forms of strabismus is still lacking.
Corsi et al.7 observed alterations in proprioceptors located at the scleral myotendinous junction of the EOMs of patients suffering from congenital strabismus. Another study20 on congenital esotropia also reported the presence of altered sensory nerve endings at the myotendinous junction, and of normal motor nerve endings in the muscle body. This damage to nerve endings consisted of alterations to both contractile structures and mitochondria, and resulted in severer lesions at the myotendinous junction than in the muscle body.20 In patients with congenital nystagmus, the myotendinous and tendinoscleral area of EOMs showed not only anomalous IMCs but also anomalous vascular endothelial cells.33 All of these alterations support the hypothesis that the most important EOM functional alteration in congenital oculomotor disorders concerns the distal myotendinous junction, and that IMCs, as proprioceptors, play a prominent role in disease pathogenesis.
In this study, we observed that almost normal features in the nerve endings at the myotendinous junction are demonstrated in acquired strabismus, including constant exotropia, intermittent exotropia, paralytic strabismus, sensory strabismus, and secondary esotropia after retinal reattachment. However it is important to note that some IMCs in noninfantile strabismus exhibited the morphologic alterations, which should be interpreted as rather acquired abnormalities from strabismus than the causes of oculomotor disorders. These findings provide evidence that IMCs have effector properties.
Based on morphological structures, it appears that congenital and early-onset oculomotor disorder may be related to a degeneration and/or dysgenesis of IMCs. Furthermore, when considering the normal features of most IMCs in acquired strabismus our study supports the hypothesis that some disturbance of IMCs may play an important role in the pathogenesis of oculomotor disorder, and that this is not the consequence of strabismus due to other causes. We speculate that IMCs are supposed to provide proprioceptive information, and a proprioceptive feedback system should be stimulated and calibrated early in life for the development of binocular vision. Our study provides evidence that some disturbances in the proprioceptive feedback network, such as degeneration and/or dysgenesis of IMCs, could result in oculomotor disorders. This concept is supported by the findings of previous studies,34,35 in which neonatal de-efferentation in the rat resulted in a reduced size of proprioceptors, whereas de-afferentation in young rats caused the formation of receptors that maintained their general architecture, but completely lacked the nerve component.
In conclusion, IMCs are supposed to function as proprioceptors, although they may have also effector properties. It should be emphasized in strabismus surgery that IMCs, as ocular proprioceptors, are located in the myotendinous region.
Notes
This work was supported by the Korea Research Foundation Grant funded by the Korean Government (MOEHRD, Basic Promotion Fund) (KRF-2007-E-00084).